

Organized Selfishness: Reframing the Gene Through Entropy.


I recently completed reading The Selfish Gene by Richard Dawkins. During my reading, I distinctly remembered the passage attached in the image. It stands out strikingly, as it demonstrates that Dawkins understands probability and systems chemistry so succinctly—that he managed to describe a very complicated process in just three sections of prose. I praise his capabilities; however, this phenomenon deserved more elaboration due to its importance in evolutionary biology and its role in establishing the foundation of The Selfish Gene.
Here, Dawkins introduces the audience to the behavior of energy and matter, illustrating how these forces can dramatically accelerate the emergence of complex structures. We could divide the process he describes into three parts: energy, molecular dynamics, and entropy. I could write ad infinitum about how each of these components influences the overall explanation; however, I will be focusing specifically on the thermodynamics of entropy.
The second law of thermodynamics was long perceived as a contradiction to understanding the persistence and emergence of complex ordered systems like living organisms. The law states that the total entropy of a closed system must increase over time, leading to a natural progression towards disorder. Deductive reasoning suggests life is defying this principle. Living systems and our components exhibit robotic levels of organization, but this is not a quasi-mystical trick, magic or the work of an intelligent being. Schrodinger solved this dilemma by turning to nonequilibrium thermodynamics for modelling living systems, that is, he recognized that living systems exist in a world of energy and material fluxes.
Consider the early prebiotic Earth, before life arose, eventually giving rise to living systems and ecosystems that function as an open thermodynamic system. “Open” meaning they perpetually exchange both energy and matter with their environment. Examples of energy fluxes entering the system may include solar radiation, geothermal heat, and lightning, all of which contribute to establishing a concept central to this framework-external energy gradients. The concept of a gradient can be described by—a difference in energy, temperature, concentration, or other physical properties across space. In the case of ecosystems, the primary gradient is the energy differential imposed by solar radiation. The Sun provides a continuous influx of high-energy, low-entropy photons. The Earth absorbs this energy, processes it through various biological and chemical means, and ultimately re-emits it as lower-energy, higher-entropy heat.
This leads to what can best be described as the “thermodynamic imperative”: systems that are driven far from equilibrium by external gradients will evolve structures and processes to reduce those gradients. In this context, the second law of thermodynamics implies that as systems are pushed away from equilibrium, they will exploit every available pathway to resist externally imposed gradients. In lay terms, systems will explore routes to maximize entropy production within the constraints they face. As a result, the second law of thermodynamics is conserved as the output of the resistance will be increased disorder to the surroundings of the system. A classic example of such a phenomenon is the Bénard cell. When a shallow layer of fluid is heated from below, convection patterns—ordered, hexagonal structures—spontaneously emerge. These patterns enhance heat transport, thus dissipating the imposed temperature gradient more effectively. Bénard cells are an example of dissipative structures: ordered formations that arise to degrade gradients more efficiently, not in violation of the second law, but as a consequence of it.
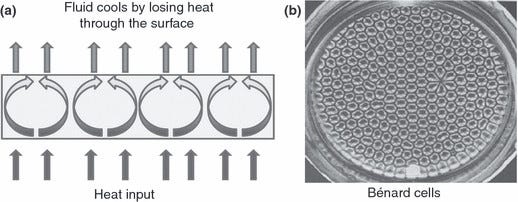
These structures self-organize as a result of a process involving tiny and random fluctuations in the system in relation to temperature or pressure, to name a few. Once such fluctuations approach a critical threshold under conditions far from equilibrium, they can grow—providing a starting point for new complex structures to emerge. The system contains small instabilities that, under current conditions, lead it to become unstable. This fuels the fluctuations to grow, and the system can no longer spring back into its original state. A key concept here is that once the previous state is unreachable, a new organization must emerge, and it is at this bifurcation point that the system effectively "chooses" a new trajectory that is irreversible. This new structure represents the selfish need to more effectively dissipate the applied gradient. The new state will therefore represent an organized, self-sustaining structure that processes energy more effectively under the current conditions. It is evident that the second law of thermodynamics in open dynamic systems provides intuitive mechanisms that explain how natural order must emerge to minimize internal entropy. The key takeaway here is for the reader to begin applying these concepts to living systems and to have the insight to see that complex structures had to precipitate. The second law here, combined with the mechanism for dissipation, acts almost as a selection mechanism that significantly reduces the time required to form infinitely complex structures.
Dawkins missed the opportunity to further convince readers by introducing the core thermodynamic behavior that explains how seemingly improbable complex structures become probable over long timescales. Misjudging dissipative structures as “random” or “divinely inspired” is almost always the result of misunderstanding the deep relationship between probability and time. To the naked eye, multicellular entities and their intricate components may appear to be the result of divine providence, because—if one assumes that such complexity arises purely by chance—the probability is so low that their emergence seems to demand more time than the universe has existed. I prepose that selection through thermodynamics must help form the backbone of the Selfish Gene argument. Genes do not persist merely because they replicate, but because their replication is embedded in larger energy-processing systems that follow the second law of thermodynamics applied to dynamic systems. Genes are not only replicators but participants in dissipative structures: the carriers of information that contribute to the emergence of order by enhancing the system's ability to degrade external gradients.
It is only right to now question whether each of these structures has a layering process that leads to even more complex structures over time. The Bénard cells are extremely trivial examples once compared to the complexity of mammalian species. Firstly, dissipative structures are only stable over a finite range of external conditions. Thus, their stability is dependent on maintaining the particular flows of energy and matter that drive the system to instability. If these fluxes—the inputs and outputs that drive the system—change beyond a certain threshold, the system may become unstable again, triggering a new bifurcation or collapse. If the bifurcation leads to the system reorganizing into a new stable system state—a condition where internal order can be maintained given the prevailing external forces—these stable states will persist until disrupted by further fluctuations or environmental shifts. Deductively, we can conclude that this is a non-deterministic process—meaning we cannot precisely measure or predict future states, as the paths the systems take are dependent on the nature of the fluctuations and fluxes. This suggests that nature exhibits a tendency to resist gradients, and as a system is driven further from equilibrium, more sophisticated mechanisms will emerge to oppose the differential. The formation of complex structures and processes represents nature’s resistance—meaning that organized behaviors and patterns are not merely advantageous in supporting the origin of life, but thermodynamically driven, necessary responses.
To conclude, I cannot fathom a more exquisitely organized system than the genes that give rise to the human being. This genetic architecture is not a miraculous anomaly, but a logical consequence of the way matter and energy interact. It reframes the behavior Dawkins describes as “selfish” in a new light—what I call organized selfishness. Organisms became survival machines not through chance alone, but through the thermodynamic imperative to minimize energy gradients. Altruistic behavior and the relentless drive for propagation arise not in spite of selfishness, but because of a deeper, organizing desire embedded in nature: to maintain structure by resisting entropy. My argument here is not to undermine natural selection, but to suggest that it operates in tandem with thermodynamic principles. I believe this not only enhances the feasibility of non-creationist theories on the origin of life, but does so through a claim that is falsifiable.
